" первая часть
Физиология
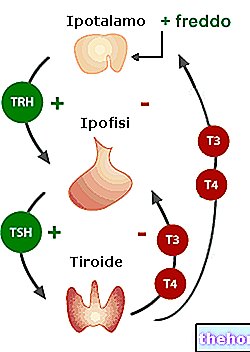
Сенсорные нейроны нервно-мышечного веретена чувствительны к растяжению.
Их промежуточная часть уже в состоянии покоя достаточно растянута, чтобы побудить их посылать нервные импульсы, которые они доверяют чувствительным волокнам. В спинном мозге эти волокна сокращают синапсы непосредственно с альфа-мотонейронами, отвечающими за иннервирование той же мышцы, от которой они начали. Благодаря этой основной активности, мышца в состоянии покоя всегда поддерживает определенную степень напряжения, определяемую как «мышечный тонус».

Во время движений веретено удлиняется и укорачивается вместе с мышцей. Следовательно, любой жест, который приводит к удлинению мышечных волокон, также воздействует на веретена таким же образом, вызывая увеличение частоты исходящих импульсов. Эти сигналы немедленно обрабатываются спинным мозгом, заставляя мышцу рефлексировать сокращение и защищая ее от повреждений, вызванных чрезмерным растяжением. Степень этого рефлекторного мышечного сокращения тем интенсивнее, чем выше частота нервных импульсов (в свою очередь, прямо пропорциональна степени растяжения, воспринимаемого сенсорными волокнами нервно-мышечного веретена).
Одновременно с активацией альфа-мотонейронов тормозные сенсорные волокна активируют тормозные интернейроны, ответственные за временное «молчание» альфа-мотонейронов, которые иннервируют мышцы-антагонисты, предотвращая их сокращение.
Все это происходит с помощью непроизвольного механизма, называемого рефлексом растяжения или миотатическим рефлексом [da мио = мышца е тасис = растяжение].
Роль гамма-мотонейронов теперь еще предстоит объяснить. Их задача - регулировать чувствительность нервно-мышечных веретен в зависимости от степени растяжения, чтобы они оставались активными даже при укорочении мышцы. Все это стало возможным благодаря так называемой альфа-гамма-коактивации, то есть одновременному сокращению фузальных и интрафузальных волокон. Поскольку последние иннервируются на обоих концах мышц, их сокращение приводит к удлинению центральной области, что удерживает сенсорные окончания в растяжении.
В то время как мышца, иннервируемая альфа-мотонейроном, укорачивается за счет снижения напряжения на капсуле веретена, одновременная активация интрафузальных волокон, гарантируемая коактивацией гамма-излучения, поддерживает нервно-мышечное веретено в активном состоянии. Таким образом, чувствительность рецептора может оставаться постоянной на каждом уровне сокращения, обеспечивая большую плавность движений и более быструю реакцию мышц в случае необходимости.
Дальнейшее исследование активности нервно-мышечных веретен предусматривает разделение волокон ядра мешочка на два подкласса: статические волокна мешка и подкласс динамических волокон мешочка. Последние, иннервируемые сенсорными волокнами типа Ia, в основном получают резкие и быстрые изменения скорости. благодаря также «двигательной иннервации, состоящей из особенно быстрых гамма-волокон (богато миелинизированных, называемых динамическими гамма-волокнами).
В то время как только что описанные волокна идеально подходят для быстрого считывания изменений длины мышцы, волокна статического мешочка предоставляют более точную информацию о продолжительности и степени изменения напряжения (они иннервируются статическими волокнами гамма II). Разряд вторичных волокон, ЦНС получает информация о «степени» удлинения мышц и, через основные, о скорости удлинения.
Наконец, следует отметить, что на активность нервно-мышечных веретен влияют несколько экстрафузальных факторов, таких как температура (тепло снижает их активность, вызывая расслабление, в то время как холод увеличивает жесткость) и степень утомления (эффективность миотатического рефлекса). снижается, когда спортсмен устает, что предрасполагает его к риску травмы мышц).