«Первые шаги гликолиза
Первый фермент, используемый во второй фазе гликолиза, - это глицеральдегид-3-фосфатдегидрогеназа; дегидрогеназы - это ферменты, которые катализируют передачу восстанавливающей силы от восстанавливающей молекулы, которая окисляется, к другой восстанавливающейся молекуле (окислительно-восстановительная реакция). Субстратами этого фермента являются НАД (никотидамид адениндинуклеотид) и FAD (флавинадениндинуклеотид).
На этой стадии дегидрогеназа катализирует превращение глицеральдегид-3-фосфата в 1,3-бисфосфоглицерат: на том же каталитическом сайте альдегидная группа окисляется до карбоксильной с последующим восстановлением NAD + до NADH, и впоследствии карбоксильная группа может с образованием ангидридной связи с ортофосфатом. Первый процесс очень экзэргонический (высвобождает энергию), а второй очень экзэргонический (требует энергии); если бы не было каталитического центра, глобальная реакция не имела бы места: первая реакция происходила бы с выделением энергии, которая рассеивалась бы в виде тепла и, следовательно, не могла бы использоваться для образования ангидридной связи.

После образования 1,3-бисфосфоглицерата фермент восстанавливает свою исходную структуру и готов действовать на новом субстрате.

Затем идет фосфоглицераткиназа который позволяет переносить фосфорил от 1,3-бисфосфоглицерата к молекуле АДФ; мы получили АТФ (один АТФ на каждую молекулу глицеральдегид-3-фосфата, следовательно, два АТФ на каждую исходную молекулу глюкозы), который компенсирует расход энергии первая фаза гликолиза.
Анион арсената (AsO43-) влияет на гликолитический путь, поскольку он может заменять фосфат в первой реакции второй фазы гликолиза, давая 1-арсенио-3-фосфоглицерат, который очень нестабилен и, как только он свободен от каталитического сайт, гидролизует, высвобождая "арсенат", возвращающийся в кровоток. Следовательно, арсенат имитирует действие фосфата и входит в каталитический сайт: в присутствии арсената реакция, которая производит АТФ (от 1,3-бисфосфоглицерата до 3-фосфоглицерата), не происходит, потому что 3-фосфатглицеральдегид преобразуется непосредственно в 3-фосфоглицерат; при отсутствии АТФ клетки погибают (отравление мышьяковой кислотой).
В третьей реакции окислительной фазы 3-фосфоглицерат превращается в 2-фосфоглицерат под действием фосфоглицератмутаза; в реакции участвует промежуточное соединение 2,3-бисфосфоглицерат.

На следующем этапе вмешивается фермент. энолаза который способен катализировать удаление молекулы воды из углеродного скелета 2-фосфоглицерата с получением пирированного фосфоенола (PEP);

PEP обладает высоким потенциалом передачи фосфорила: он переносится под действием фермента пируваткиназа, фосфорил к АДФ с образованием АТФ на пятой стадии второй фазы с получением пирувата.

2-фосфоглицерат и 3-фосфоглицерат обладают низкой способностью переноса фосфорила, поэтому для получения АТФ из этих молекул 3-фосфоглицерат превращается в 2-фосфоглицерат во время гликолиза, поскольку он получается из последнего. виды с высоким потенциалом переноса.
Прежде чем продолжить, позвольте открыть скобки для 2,3-бисфосфоглицерата; последний присутствует во всех клетках, в которых гликолиз происходит в очень низкой концентрации (это промежуточное звено третьей реакции второй фазы гликолиза). В эритроцитах, с другой стороны, 2,3-бисфосфоглицерат имеет постоянную концентрацию 4-5 мМ (максимальная концентрация), потому что они обладают ферментативным наследием, задачей которого является его производство; в эритроцитах наблюдается отклонение от гликолиза с образованием 2,3-бисфосфоглицерата: 1,3-бисфосфоглицерат превращается в 2,3-бисфосфоглицерат под действием бисфосфоглицератмутаза (эритроцит) и 2,3-бисфосфоглицерат под действием бисфосфоглицерат фосфатаза (эритроцит) становится 3-фосфоглицератом. Затем в эритроцитах часть 1,3-бисфосфоглицерата, полученного в результате гликолиза, превращается в 2,3-бисфосфоглицерат, который затем возвращается в гликолитический путь в виде 3-фосфоглицерата; при этом происходит третья стадия окислительной фазы гликолиз, в результате которого получают АТФ. Количество потерянного АТФ - это цена, которую эритроцит готов заплатить за поддержание концентрации 2,3-бисфосфоглицерата, в которой нуждаются эти клетки, потому что это влияет на способность «гемоглобина связывать» кислород.
Мы видели, что в первой реакции второй фазы гликолиза НАД + восстанавливается до НАДН, но необходимо, чтобы после получения пирувата НАДН снова превращался в НАД +: это происходит при молочной ферментации (получается лактат). или путем спиртовой ферментации (в игру вступают пируватдекарбоксилаза, декарбоксилатирующая пируват, и дегидрогеназа, которая образует этанол); в ферментациях не участвует кислород (анаэробы).
Из-за молочной ферментации молочная кислота, если ее не утилизировать должным образом, накапливается в мышцах и, высвобождая H +, вызывает непроизвольное сокращение мышц и, следовательно, судороги; мышца при сильном стрессе также может достигать минимального pH 6,8.
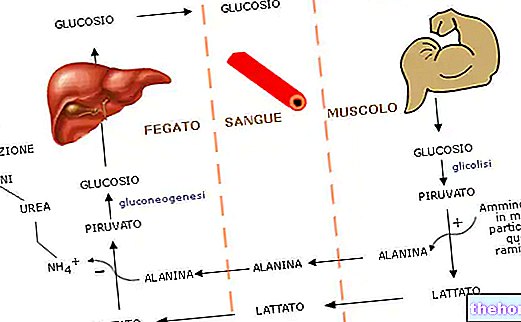
Через цикл Кори часть мышечной усталости передается на печень, когда мышца перегружена. Предположим, что мышца работает без снабжения кислородом (ошибочное предположение): если мышца работает умеренно, АТФ, необходимый для сокращения, обеспечивается исключительно за счет гликолиза. Если активность мышцы увеличивается и требуется дополнительный АТФ, ускорьте аэробный метаболизм, преобразовывая лактат, который таким образом удаляется, превращается в глюкозу. В действительности, мышца задействует аэробный метаболизм: при наличии кислорода мышца использует, прежде всего, АТФ, обеспечиваемый аэробным метаболизмом, а при отсутствии кислорода анаэробный метаболизм ускоряется за счет цикла Кори. Этот цикл предполагает, что лактат передается из мышцы в печень, где, расходуя энергию, вырабатывается больше глюкозы, которая возвращается в мышцу. В ходе этого цикла часть АТФ, потребляемого в мышце, доставляется печенью, которая: в процессе глюконеогенеза способен производить глюкозу, которая может использоваться мышцами для получения АТФ.
Метаболизм глюкозы, описанный до сих пор, не включает кислород, но аэробный метаболизм глюкозы позволяет получать в 17-18 раз большее количество АТФ, чем полученное с помощью гликолитического пути, поэтому, когда клетка имеет возможность выбирать между аэробным и эд анаэроб, отдает предпочтение первому.
При аэробном метаболизме пируват попадает в митохондрии, где он претерпевает преобразования и в конечном итоге образуется диоксид углерода и вода; таким образом на каждую молекулу деградированной глюкозы получают 34 молекулы АТФ.


.jpg)